Среди множества биологических объектов, к исследованию которых применялись методы зондовой микроскопии, первой была молекула дезоксирибонуклеиновой кислоты. Однако на начальном этапе этих исследований отмечались сложности в интерпретации получаемых изображений. Так, авторы работ [1,2] представили данные сканирующей туннельной микроскопии молекул ДНК, адсорбированных на подложку из высокоориентированного пиролитического графита (пирографита). Выбор данного материала в качестве подложки был обусловлен его проводящими свойствами, а также простотой приготовления атомарно гладких участков поверхности значительной площади. На СТМ-изображениях наблюдались протяженные цепочки, обладающие продольной периодичностью (период около 2-3 нм), на основании чего авторы делали вывод о визуализации витков двойной спирали.
Однако позднее были опубликованы данные [3], свидетельствующие о возможности наблюдения сходных ``ДНК-подобных'' структур при исследовании методом СТМ чистой, свежесколотой поверхности пирографита. Возникновение таких структур объясняется чувствительностью СТМ к электронным свойствам поверхности - происходит визуализация доменных стенок пирографита [4]. Эти результаты, а также слабая воспроизводимость и отсутствие безусловно необходимых контрольных экспериментов в первых работах по СТМ-визуализации ДНК вызывали сомнения в применимости методов зондовой микроскопии для исследования макромолекул.
Существенное изменение ситуации произошло в 1992 г., когда были опубликованы первые надежные и воспроизводимые результаты исследования ДНК методом атомно-силовой микроскопии (АСМ) [5,6]. Таким образом, прошло более пяти лет с момента создания атомно-силового микроскопа (1986 г.) и более десяти лет существования зондовой микроскопии, прежде чем был достигнут определенный успех в применении метода к исследованию биологических объектов. В то же время методы зондовой микроскопии весьма успешно применялись в течение этих десяти лет для исследований структуры поверхности твердых (кристаллических) тел. За эти годы возникло понимание, что потенциал возможностей зондовой микроскопии в исследованиях биологических объектов может быть реализован лишь при решении в каждом конкретном случае основной экспериментальной задачи: определения адекватной методики препарирования образцов.
Универсальной методики приготовления образцов для решения широкого спектра задач зондовой микроскопии пока не существует, поэтому используемые методики определяются спецификой задачи, т.е. являются методиками ``на конкретный случай''. Применительно к исследованиям нуклеиновых кислот во многих случаях бывает важно, чтобы макромолекулы адсорбировались на подложку в расправленном состоянии. Для успешного исследования необходимо также, чтобы исследуемые молекулы достаточно прочно фиксировались на подложке. В противном случае, при исследованиях в жидкой среде, молекулы будут увлекаться зондом в процессе сканирования; при исследовании на воздухе, силы, действующие на молекулу при высыхании капли препарата на подложке, также будут увлекать адсорбируемые структуры, приводя к неоднородному (с образованием агрегатов) поверхностному распределению исследуемых объектов. Ниже рассмотрим основные методики, позволяющие зафиксировать молекулы нуклеиновых кислот на поверхности в расправленном состоянии.
Первые надежные результаты исследования ДНК методом атомно-силовой микроскопии были получены в том случае, когда изучаемые структуры наносили из капли рабочего раствора (раствора буфера, водно-спиртовой среды и т.п.) на поверхность слюды, модифицированной ионами двух- (и более) валентных металлов [5,6]. Эти ионы, по-видимому, служат связующими мостиками между отрицательно заряженной слюдой (в водных растворах) и отрицательно заряженными фосфатными группами молекулы ДНК. Модификация слюды может предшествовать процессу адсорбции макромолекул - в этом случае свежесколотую слюду помещают для предварительной обработки на некоторое время (минуты, часы) в раствор, содержащий катионы металлов, затем промывают (водой, водным раствором этанола и пр.) и высушивают. После этого на модифицированную поверхность наносят рабочий раствор с исследуемыми структурами. Другой подход - добавление солей металлов непосредственно к рабочему раствору - позволяет исключить этап предварительной обработки слюды (препарат наносят непосредственно на поверхность свежего скола). После высыхания капли рабочего раствора образцы, как правило, подвергают дополнительной промывке.
В работе [7] методом АСМ проводили сравнительный анализ процесса адсорбции макромолекул ДНК на слюду из раствора, как с катионами металлов, так и без них. Было показано, что во втором случае адсорбированные молекулы нестабильны в процессе промывки образцов и при сканировании. Кроме того, АСМ-изображения макромолекул характеризуются большим количеством запутанных и перекрученных участков, тогда как при присутствии в растворе катионов металлов и применении предшествующей сканированию процедуры промывки образцов молекулы ДНК фиксируются на поверхности подложки в расправленном состоянии.
Авторы работы [8] исследовали методом АСМ в жидкостной ячейке в реальном масштабе времени обратимое осаждение молекул ДНК на слюду при различных параметрах рабочего раствора. Было показано, что адсорбция объектов на поверхность подложки и величина силы адгезии существенно зависят от концентрации связующих катионов (оптимальное значение около 2 мМ для Zn 2+) и pH среды (оптимальное значение около 7,5).
На сегодняшний день использование катионов металлов для связывания ДНК с подложкой является, пожалуй, наиболее распространенной методикой приготовления образцов при АСМ-исследованиях, что объясняется ее простотой и высокой воспроизводимостью результатов.
На рис. 1.1 представлены АСМ-изображения молекул ДНК, адсорбированных на поверхность слюды в присутствии катионов Ni 2+ (а) или уранилацетата (б). Из рисунка видно, что молекулы зафиксированы на подложке в достаточно расправленном состоянии.
Приводимые изображения молекул характеризуются завышенными латеральными размерами - 10-15 нм - и заниженными вертикальными - около 0,7 нм, в то время как диаметр одиночной молекулы ДНК около 2 нм. Завышение латеральных размеров полученных изображений обусловлено конечной величиной радиуса кривизны зонда (количественная методика учета данного артефакта и определения реальной ширины объекта разработана в разделе 2.2). Причиной занижения вертикальных размеров может являться деформация молекулы под действием зонда (рассмотрение теории контактных деформаций и применение ее для описания эффекта занижения высот АСМ-профиля объекта приведено в разделе #.#). Следует подчеркнуть, что эти артефакты являются типичными при АСМ-исследованиях молекул нуклеиновых кислот [9].
Другая широко используемая методика химической модификации подложки для стабилизации молекул нуклеиновых кислот на поверхности - это силанизация слюды [10]. Процесс силанизации существенно не изменяет топографических особенностей поверхности слюды - она остается достаточно гладкой, в то же время улучшается связь макромолекулы с модифицированной подложкой. В работе [11] при проведении АСМ-исследований нуклеиновых кислот (ДНК, двунитевой рибонуклеиновой кислоты (РНК)) в качестве подложки использовали поверхность слюды, химически модифицированную 3-аминопропилтриэтоксисиланом (APTES). Модификация включала ковалентную привязку аминогрупп молекул APTES к слюде, степень модификации контролировали с помощью АСМ, и выбирали таким образом, чтобы шероховатость получаемой поверхности не затрудняла идентификацию исследуемых структур. Это обстоятельство (необходимость контроля степени шероховатости модифицированной подложки) незначительно усложняет методику.
Было показано [12], что этот подход препарирования образцов достаточно эффективен в рутинных исследованиях распределения молекул нуклеиновых кислот по длинам. Представленные авторами результаты совпадали с данными электронной микроскопии (исследовались молекулы двунитевой РНК ретровируса), при этом было отмечено, что при равенстве разрешающих способностей важным преимуществом АСМ является существенно менее сложная методика приготовления образцов.
Была продемонстрирована возможность использования модифицированной APTES слюды при проведении исследований как на воздухе, так и в водной среде - в обоих случаях величина адгезии молекул нуклеиновых кислот к подложке позволяла осуществлять сканирование в контактном режиме. В работе [13] осуществляли сканирование одного и того же участка поверхности с адсорбированными макромолекулами на воздухе и в воде, при этом во втором случае отмечалось увеличение разрешающей способности (примерно в три раза), что может быть связано с исключением негативного влияния капиллярных сил (см. раздел #.#.#). Капиллярные силы увеличивают силовое воздействие зонда на образец, что вызывает дополнительные деформации макромолекулы, ее разрушение или нестабильность в процессе сканирования, и приводит к уменьшению разрешающей способности.
Другие исследователи также сообщают, что, если АСМ-исследования проводятся либо в сухой газовой атмосфере (иногда для уменьшения относительной влажности температуру рабочей атмосферы повышают до 60-100°С [14]), либо в жидкостной ячейке [15,16], то влияние капиллярных сил уменьшается, что увеличивает достигаемое пространственное разрешение. Применение режима прерывистого контакта (tapping mode, см. раздел #.#.#) [17,18] также позволяет исключить влияние капиллярных сил; микроскопия прерывистого контакта позволяет сегодня достичь разрешения (при проведении исследований в жидких средах), необходимого для визуализации витков двойной спирали ДНК [19].
На пути подбора адекватного подхода к приготовлению образцов может оказаться полезным накопленный за десятилетия исследований багаж методик препарирования образцов электронной микроскопии. В упрощенном варианте многие из этих методик могут применяться и для решения задач зондовой микроскопии. Так, модифицированный метод Кляйншмидта [20,21] (метод белковой пленки, см. раздел 1.2) дает хорошие результаты при использовании гидрофобных подложек [22]. Недостатком метода может служить то, что комплексообразование молекул нуклеиновых кислот с белковой пленкой (часто используют пленку цитохромов C) затрудняет исследование взаимодействия этих молекул с другими белками, что ограничивает круг решаемых задач.
Для фиксации развернутых молекул ДНК на поверхности подложки применяют также бензилдиметилалкиламмоний хлорид (BAC) [23,24] - широко используемый в электронной микроскопии реагент, представляющий собой катионное поверхностно-активное вещество (ПАВ). BAC в малых концентрациях (около 5×10-5%) добавляют непосредственно в раствор, содержащий макромолекулы, перед нанесением препарата на поверхность слюды. Вследствие свойств ПАВ, на поверхности капельки, наносимой на подложку, формируется стабильная пленка молекул BAC (со связанными с ней молекулами ДНК), которая распределяется по поверхности слюды и стабилизируется за счет сил электростатического взаимодействия. Использование других ПАВ (в малых концентрациях) также способствует стабилизации молекул нуклеиновых кислот на поверхности слюды. Так, в работе [25] для разворачивания ДНК на поверхности, успешно применили 2,4,6-трис(диметиламинометил)фенол (DMP-30) и хлорид цетилпиридиния (CP), представляющие собой, соответственно, неионное и катионное ПАВ. На рис. 3.1 представлен результат АСМ-исследования ДНК бактериофага Т4, адсорбированной на поверхность слюды из раствора, содержащего малые концентрации цетилтриметиламмоний бромида (ЦТАБ).
К недостаткам метода с применением ПАВ можно отнести некоторую поверхностную неоднородность получаемых образцов, что обусловлено неконтролируемостью процессов формирования пленки и комплексообразования с исследуемыми макромолекулами. Удачным дополнением метода может служить использование методики Ленгмюра-Блоджетт: формирование на границе раздела фаз жидкость/газ пленки амфифильных молекул и последующее контролируемое перенесение ее на поверхность твердой подложки (см. главу #).
В работе [26] для иммобилизации молекул ДНК применили другую методику, сходную с традиционной для электронной микроскопии. Макромолекулы адсорбировали на поверхность слюды, сверху напыляли слой углерода. После этого на углерод приклеивали стальную пластинку, слюду удаляли, и открытой для сканирования оказывалась внутренняя (обращенная прежде к слюде) поверхность углеродной пленки с жестко закрепленными в ней макромолекулами. Шероховатость поверхности углеродной пленки составляла, по оценкам авторов, единицы ангстрем и допускала однозначную идентификацию отдельных молекул ДНК. Авторы работы отмечают высокую стабильность приготовленных образцов - макромолекулы не разрушались при больших силах воздействия зонда, результаты не зависели от среды исследования (эксперименты проводились как на воздухе, так и в жидкости), длительное хранение не влияло на качество образцов. Недостаток этого метода состоит в его относительной экспериментальной трудоемкости.
Успехи применения атомно-силовой микроскопии к исследованию молекул нуклеиновых кислот позволили достичь прогресса и при проведении подобных экспериментов методами сканирующей туннельной микроскопии. Хорошие результаты были получены при СТМ-исследовании молекул ДНК, адсорбированных на химически модифицированных поверхностях металлов [27]. Химическая модификация включала ковалентную связь химически поляризуемых групп тиолов с чистой поверхностью металла. Адсорбция ДНК осуществлялась за счет кулоновского взаимодействия молекулы с плотно упакованной мономолекулярной пленкой тиолов, ориентированной положительными функциональными группами к молекуле ДНК. Характерной особенностью представленных в работе [28] изображений молекул ДНК (приготовленных по описываемой методике) является отрицательный контраст (при движении над молекулой туннельная игла опускается ниже, чем при движении над поверхностью подложки), связанный, по-видимому, со слабой проводимость макромолекулы.
Другой перспективный подход к исследованию макромолекул методами СТМ - применение низкотоковой туннельной микроскопии [29] с рабочим диапазоном туннельного тока менее 1 пА. В этом случае экспериментальные результаты [30] свидетельствуют о возможности визуализации молекул ДНК, адсорбированных на поверхности диэлектрической подложки (слюды). Транспорт заряда в системе зонд-образец-подложка осуществляется за счет проводимости ультратонкой пленки воды, покрывающей поверхности образца и подложки при проведении исследований в условиях контролируемой относительной влажности. Оптимальное значение последней для подобных экспериментов составляет 60- 65%.
Т.о. спектр применяемых методик препарирования образцов нуклеиновых кислот для СЗМ весьма широк. В то же время стоит отметить, что описанные методики разработаны для исследования свободных молекул ДНК, поэтому априори неизвестно адекватны ли они для изучения, например, взаимодействия нуклеиновых кислот с белками или поверхностно-активными веществами. Упомянем также, что в литературе по СЗМ описаны, в основном, лишь исследования двунитевых молекул нуклеиновых кислот: РНК и ДНК, имеющих двойную спираль, и практически отсутствуют данные об исследовании однонитевых молекул (которые сложнее зафиксировать на подложке в расправленном состоянии), см. главу 2.
При подборе адекватной методики приготовления образцов сканирующий зондовый микроскоп представляет собой удобный и надежный прибор для исследования свойств биологических структур на молекулярном уровне. Убедительным доказательством этого утверждения могут служить результаты работ, посвященных исследованию конформационных свойств молекул нуклеиновых кислот и их комплексов с белками, ПАВ и т.п.
Авторы работы [31] исследовали влияние специфических лигандов (дистамицина и микрогонотропена-6b) на конформационные свойства нескольких типов молекул ДНК методом АСМ прерывистого контакта. Результаты исследований не только обеспечили визуальное доказательство влияния данных лигандов на конформацию ДНК, но и позволили сделать определенные количественные оценки, основанные на анализе представленных авторами гистограмм распределения расстояния между концами ДНК.
В работе [32] исследовали специфическое взаимодействие РНК-полимеразы из Escherihia coli с ДНК, содержащей lPL промотор. Образованные комплексы были визуализированы методом АСМ, на полученных изображениях наблюдались молекулы ДНК с РНК-полимеразой, локализованной на расстоянии 4/9 от одного из концов. Проводили анализ структуры комплекса РНК-полимеразы и ДНК как в открытых комплексах, связанных с промотором (OPCs), так и в стабильных элонгационных комплексах (С15) с образованным 15-нуклеотидным транскриптом. Было показано, что в области прикрепления РНК-полимеразы происходит изгиб молекулы ДНК, причем средний угол изгиба для OPCs и С15 составлял 54±31° и 92±37°, соответственно. На основании обнаруженного различия в значениях средних углов изгиба для OPCs и C15 авторы делают вывод, что процесс транскрипции сопровождается изменением структуры комплекса РНК-полимеразы с ДНК, обусловленной конформационными изменениями полимеразы.
Возможность АСМ-визуализации молекулярных процессов продемонстрирована в работе [33] на основе последовательного анализа образования неспецифичных комплексов молекулы ДНК и РНК-полимеразы в реальном масштабе времени. Схема эксперимента включала в себя адсорбцию макромолекул ДНК на слюду из 10 мM HEPES буфера в присутствии ионов магния (1-10 мM MgCl2), затем следовали промывка и высушивание образцов в эксикаторе. После этого образцы помещали в жидкостную ячейку микроскопа, начинали процесс сканирования, а затем в раствор вводили РНК-полимеразу. Было показано, что образование комплексов белок-ДНК наблюдается уже через несколько секунд после добавления полимеразы, что свидетельствует о сохранении нативной конформации РНК-полимеразы и ДНК в процессе приготовления образцов и проведения исследований. Эти результаты демонстрируют возможность использования зондовой микроскопии для исследования процессов, ответственных за распознавание и сборку макромолекулярных комплексов в физиологических условиях.
Большое прикладное значение имеет возможность решения методом АСМ задачи физического картирования ДНК: специфические участки макромолекулы помечаются определенными маркерами, анализ местоположения этих маркеров на получаемых АСМ-изображениях позволяет составлять карты относительного расположения специфических последовательностей нуклеотидов. Так, в работе [9] продемонстрирована возможность визуализации местоположения биотина, ковалентно привязанного к первому нуклеотиду праймера. Биотин помечался белковым комплексом стрептавидин-стафиллококковый белок А (стрептавидин имеет высокую способность связывания с биотином, а белок А увеличивает размеры маркера для однозначной его идентификации на АСМ-изображениях).
В работах [34,35] был использован метод картирования клонированных в плазмидный вектор последовательностей LTR (длинные концевые повторы) с помощью специфических маркеров, имеющих характерную форму - R-петель. R-петли формировались последовательностями РНК из 345 и 380 нуклеотидных оснований, комплементарными к U3 и U5 областям LTR человеческого эндогенного ретровируса K-10 (HERV-K10). В процессе образования петли РНК формировала двойную спираль с комплементарным участком ДНК, вытесняя вторую нить ДНК, которая коллапсировала в результате воздействия ионов Mg 2+; характерная форма образованной структуры (петля) позволяла уверенной идентифицировать ее на АСМ-изображениях. Различная длина зондов позволяла уже из одной гистограммы определить как положение, так и ориентацию LTR. Полученные результаты позволяют предположить, что в будущем разрешение приборов СЗМ может быть достаточным для определения последовательности нуклеотидов ДНК.
Уникальную возможность зондового микроскопа как прибора, позволяющего проводить прецизионные исследования локальных свойств поверхности, продемонстрировали авторы работы [36]. Они проводили прямые исследования силового взаимодействия, ответственного за формирование витков молекулы ДНК, по следующей схеме.
На поверхностях подложки и кремниевого микрозонда создавали два типа покрытия, представляющего собой слой ковалентно привязанных за один из концов комплементарных олигомеров - в одном случае (АЦТГ)5, в другом (ЦАГТ)5. В процессе взаимодействия пары данных олигонуклеотидов (длиной в 20 нуклеотидов) возможно образование комплексов с 20, 16, 12, 8 и 4 парами взаимодействующих оснований.
В ходе эксперимента многократно измеряли кривую силового взаимодействия F(z) между зондом и подложкой. На основании этих результатов определяли силу адгезии и строили гистограммы ее распределения. В случае отсутствия специфического взаимодействия между комплементарными участками наблюдалось бы однородное гауссово распределение для силы адгезии. Однако на полученных гистограммах четко прослеживались отдельные пики (1,52±0,19; 1,1±0,13; 0,83±0,11 нН), отображающие, очевидно, специфическое взаимодействие 20, 16 и 12 пар комплементарных оснований (авторы указывают, что комплексы с 8 и 4 взаимодействующими парами оснований термодинамически нестабильны при 27°С - температуре проведения исследований).
Сходную экспериментальную схему - с образованием между взаимодействующими поверхностями ``мостиков'' в виде отдельных нитей ДНК длиной в 160 оснований - использовали для анализа внутримолекулярных упругих свойств. Таким образом, авторам удалось продемонстрировать возможность применения АСМ для прямого количественного анализа межмолекулярного и внутримолекулярного взаимодействия в комплексах биологических и синтетических макромолекул, что открывает новые перспективы для обнаружения и локализации специфических последовательностей нуклеотидов с ангстремным разрешением.
В настоящее время в арсенале исследователей, изучающих биологические и органические структуры, имеется широкий спектр различных физических методов исследования. Для того чтобы прояснить возможности и перспективы метода сканирующей зондовой микроскопии и обозначить ее место в ряду других методов исследования, ниже приведена краткая характеристика основных экспериментальных подходов к исследованию биологических и органических структур.
К дифракционным относятся методы реконструкции реальной структуры объекта путем анализа дифракции взаимодействующего с ним пучка электронов, нейтронов или рентгеновский лучей: математическая обработка дифракционной картины позволяет решить обратную задачу и восстановить структуру объекта. Принципиальная трудность метода заключается в невозможности получения непосредственно из эксперимента информации о фазах дифрагировавших лучей. Один из возможных путей решения данной проблемы - использование меток - атомов тяжелых металлов, присоединяемых к определенным группам исследуемого объекта. Так, последовательное мечение различных специфических групп гемоглобина позволило получить необходимую информацию для расчета структуры этого белка [37] по данным рентгеноструктурного анализа его кристаллов. Этот метод нашел применение и при исследовании некристаллических (например, волокнистых) структур. Именно по косвенным рентгеноструктурным данным Уотсон и Крик впервые определили для молекулы ДНК диаметр и параметры спирали (наличие 10 эквивалентных групп в одном обороте, равном 3,4 нм). На основании этих результатов ими и была впервые предложена общепринятая ныне модель строения молекулы ДНК [38].
Дифракционные методы успешно используются и для анализа свойств поверхности, позволяя достигать высокой точности в определении реконструируемых пространственных параметров (при условии достаточной упорядоченности исследуемой структуры). Особенно широко используется метод электронографии, поскольку, из-за сильного взаимодействия электронов с веществом, определяющим в дифракционной картине является вклад именно ближайших к поверхности атомных слоев. В случае рентгеноструктурных исследований поверхности приходится использовать ``скользящую'' геометрию падающего луча и достаточно мощный источник излучения.
Метод рентгеноструктурного анализа применим и для исследований систем частиц, расположение которых не характеризуется ближним или дальним порядком: в этом случае анализируют рассеяние рентгеновских лучей под малыми углами. Этот подход позволяет определять либо форму, либо размеры микрообъектов системы (для решения задачи какая-то информация должна быть известна [39]). Если объекты исследуемого ансамбля характеризуются некоторым распределением по параметру, через который выражается искомый, то точность его восстановления ограничена. Например, при восстановлении формы частиц, разброс их по размерам резко снижает точность решения обратной задачи (если оно вообще может быть найдено) [39], и наоборот. Дополнительная информация, необходимая для корректной постановки обратной задачи, может быть получена с помощью других методов исследования - в том числе сканирующей зондовой микроскопии.
Метод электронной микроскопии (ЭМ), за годы своего существования, был дополнен множеством вспомогательных методик препарирования образцов (при исследовании биообъектов) [40]. Проблема в том, что биообъекты состоят из веществ с малыми атомными номерами, следствием чего является малое число рассеянных электронов и, в итоге, отсутствие разрешения. Сегодня наиболее часто используемые способы контрастирования биообъектов для ЭМ-исследований - это приготовление реплик, оттенение запылением металлами, ``окрашивание'' отдельных частиц и молекул ионами тяжелых металлов. Высокое разрешение дает предложенный Холлом [41] метод негативного контрастирования, принцип которого состоит в помещении исследуемых образцов в пленку аморфного вещества высокой плотности (т.е. хорошо рассеивающего электроны).
При исследовании нуклеиновых кислот методом ЭМ важной проблемой является нанесение исследуемых структур на подложку. При высушивании капли разбавленного раствора молекул на поверхности наиболее часто используемых в ЭМ подложек (коллодий, формвар, углерод) происходит агрегация исследуемых структур. Нанесение молекул ДНК с помощью пульверизатора приводит к их деградации. Поэтому большое распространение получила методика Кляйншмидта и Цана [20], основанная на взаимодействии нуклеиновой кислоты с основными белками и способности этих белков образовывать монослой на поверхности раздела фаз воздух/водный раствор. Молекулы нуклеиновых кислот, связываясь с белковой пленкой, переходят из состояния трехмерного клубка в двумерное расправленное состояние на поверхности водной субфазы. Белковую пленку с молекулами нуклеиновой кислоты переносят на сетку, после чего применяют стандартные методы контрастирования и осуществляют исследование [21]. Недостатком метода является то, что белковая пленка, покрывающая молекулу, увеличивает ее ``толщину'' до 5- 8 нм и затрудняет решение ряда задач (например, исследования взаимодействия нуклеиновых кислот с белками).
Существенной проблемой электронной микроскопии является большая вероятность возникновения артефактов, связанных с необходимостью использования специфических методик контрастирования и фиксирования биообъектов на подложке.
Зондовая микроскопия по общей схеме эксперимента близка ЭМ и для многих объектов позволяет достигать латерального разрешения, близкого к разрешению ЭМ. В то же время, латеральное разрешение СЗМ по нормали выше (метод запыления под углом в ЭМ не дает высокой точности в определении высот объектов из-за конечного размера зерна запыления), что позволяет успешно решать специфические задачи, весьма сложные для ЭМ: например, по увеличению высоты локального участка АСМ-изображения молекулы ДНК, детектировать местоположение триплекса в составе молекулы [42].
Оптические методы исследования получили в последнее время большое распространение (особенно с применением лазерного излучения). Среди наиболее часто используемых методик - спектрофотометрия, основанная на изменении спектра поглощения молекулы при димеризации (в общем случае - полимеризации) за счет взаимодействия соседних мономеров (в основном играет роль диполь-дипольное взаимодействие). Спектрофотометрия позволяет определять концентрации нуклеиновых кислот, определять степень спиральности и исследовать переход клубок ® глобула. Однако результаты исследования чувствительны к наличию в растворе ионов, способных взаимодействовать с исследуемыми молекулами (в том числе протонов, определяющих pH среды). Это связано с возможностью влияния подобного взаимодействия на контролируемые в процессе исследования спектральные изменения. Для выяснения природы спектральных изменений необходимы вспомогательные эксперименты.
Наряду со спектрофотометрией в исследованиях биообъектов широко применяются инфракрасная спектроскопия, спектрополяриметрия, спектрально-флуоресцентные исследования и т.д. Общей чертой данных методик является их косвенный, непрямой (относительный) характер - результатом исследования является усредненный по ансамблю отклик объектов. Проблемой является и необходимость решения обратной задачи для извлечения информации об объекте, что, в ряде случаев, может привести либо к значительной погрешности в результатах, либо к неадекватности решения.
Отдельно следует упомянуть метод флуоресцентной микроскопии, который нашел применение при визуализации одиночных макромолекул в нативных условиях и исследовании изменений их морфологии в процессе взаимодействия с различными лигандами и пр. К недостаткам метода следует отнести невысокую пространственную разрешающую способность (ограниченную пределом оптического микроскопа l/2, поэтому успешно можно исследовать лишь достаточно большие молекулы), и необходимость комплексообразования макромолекулы с молекулами флуоресцентного ``красителя''.
Широко используемые гидродинамические методы исследования биополимеров (вискозиметрия, седиментация, двойное лучепреломление в потоке) позволяют определить приближенную форму, размеры, молекулярный вес и деформируемость исследуемых молекул. Информация, получаемая с помощью гидродинамических методов, также является неабсолютной (усредненной по ансамблю), что подразумевает необходимость сравнения с результатами других методов исследования. Актуальной является проблема корректной постановки и решения обратной задачи для определения параметров объектов.
В этой связи методы сканирующей зондовой микроскопии призваны занять важное место в ряду традиционных исследовательских подходов к изучению биообъектов. Поскольку приборы зондовой микроскопии предназначены для исследования свойств поверхности, то стандартная схема СЗМ-исследования биообъектов включает использование достаточно ровной подложки (проводящей в случае СТМ), на которую осуществляется адсорбция изучаемых структур. Наиболее распространенные в сканирующей зондовой микроскопии подложки - атомарно гладкие сколы слюды, пирографита, дисульфида молибдена, а также эпитаксиально выращенные пленки металлов.
Необходимость использования подложки сближает методы СЗМ с электронной микроскопией. Основные отличия и преимущества сканирующей зондовой микроскопии в сравнении с ЭМ - возможность проведения исследований в различных средах (в том числе в жидкости), отсутствие необходимости какого-либо дополнительного контрастирования исследуемых структур, большая разрешающая способность по нормали, неразрушающий характер исследования (обусловленный невысокой интенсивностью взаимодействия зонд-образец). Т.о. процедура приготовления образцов для СЗМ проще и исследования можно проводить в условиях, близких к нативным.
Все это свидетельствует о том, что зондовая микроскопия является мощным современным методом исследования поверхности, призванным обеспечить дальнейшее развитие традиционным подходам. Результаты исследований последних лет убедительно продемонстрировали, что для решения многих задач метод зондовой микроскопии является самодостаточным (т.е. нет необходимости контрольного сравнения с результатами других исследований); к таковым задачам относятся, например, физическое картирование молекул нуклеиновых кислот [35], исследование изменения их морфологии в жидких средах, анализ их взаимодействия с белками [31,32], анализ структуры тонкопленочных покрытий (см. главу #) и пр.
Тем не менее, в процессе исследования большое внимание следует уделять возможным механизмам возникновения артефактов. Достаточно очевидным фактом является необходимость учитывать конечность размеров зонда, приводящую к завышению латеральных размеров исследуемых структур (раздел #.#), а также эффект занижения высот, связанный с контактными деформациями (раздел #.#). Кроме того, при исследовании многокомпонентных образцов, надлежит проводить дополнительный анализ каждой из компонент, с целью определения их парциального влияния на результат исследования, что позволит избежать ошибок в идентификации комплексов, полученных в результате взаимодействия исходных объектов.
Постановка адекватных контрольных экспериментов, безусловно, способствует уменьшению вероятности ошибки при интерпретации результатов СЗМ-исследований; избежать ошибки позволяет и сравнение с результатами других методов исследования (рассмотренных выше).
Вирусные частицы являются простейшими природными объектами, способными к репликации. Они состоят из носителя генетической информации (молекулы нуклеиновой кислоты) и оболочки, образованной белковыми молекулами. Репликация вирусных частиц осуществляется только внутри клеток живых организмов, при этом ее процесс регулируется спецификой взаимодействия молекулы нуклеиновой кислоты с белками.
Вирус табачной мозаики представляет собой полый цилиндр, имеющий длину около 300 нм, внутренний диаметр 4 нм и внешний диаметр 18 нм (согласно данным электронной микроскопии [40]). Каждая вирусная частица состоит из 2200 белковых субъединиц, организованных в правовинтовую спираль с шагом 16 1/3 субъединиц на виток. Вирусная РНК, состоящая из 6400ё6600 нуклеотидов, также образует спираль, каждый виток которой располагается между двумя витками спирали белковых субъединиц, см. рис. 2.1.
|
Вирус табачной мозаики является удобной модельной системой для апробирования методик приготовления образцов, позволяющих проводить АСМ-исследования взаимодействия вирусной РНК с белками.
В настоящее время разработаны различные методы фиксации макромолекул ДНК и РНК на поверхностях различных подложек для исследований СЗМ, см. раздел 1.1.1. Так, авторы работы [43] использовали традиционную методику иммобилизации нуклеиновых кислот на поверхности слюды катионами Mg2+, добавляемыми в рабочий раствор. В качестве объектов исследования они выбирали, в частности, короткие (25 и 50 оснований) молекулы однонитевой ДНК, а также РНК-гомополимера поли-A. Авторы обнаружили, что по используемой методике препарирования образцов только молекулы поли-А возможно иммобилизовать на подложке в расправленном состоянии - в отличие от адсорбированных молекул однонитевой ДНК, которые характеризовались компактной глобулярной структурой. Это наблюдение показывает, что основным препятствием при расправлении на подложке однонитевых молекул нуклеиновых кислот является образование внутримолекулярных пар оснований (по механизму Уотсона-Крика [38]). Действительно, молекулы ДНК, используемые авторами работы [43], могли формировать подобные внутримолекулярные пары (судя по приводимой авторами последовательности нуклеотидов этих молекул), в отличие от молекул поли-A, являющегося гомополимером, образованным лишь одним типом нуклеотидов.
Традиционная методика приготовления РНК в расправленном состоянии для электронной микроскопии включает образование комплекса РНК с монослойной пленкой белков (цитохром С или диизопропилфосфаттрипсин) на поверхности раздела фаз воздух/вода и последующий перенос пленки с молекулами РНК на поверхность пленки-подложки [21]. Данная методика позволяет получать ЭМ-изображения молекул РНК, находящихся в двумерном расправленном состоянии [44], однако в этом случае тот факт, что РНК находится в комплексе с белками (в белковой ``шубе''), может ограничить решение задач, связанных с тонкой структурой РНК и с исследованиями РНК-белкового взаимодействия.
Наблюдение свободной РНК позволяет проводить как физическое картирование элементов вторичной структуры РНК, так и исследовать специфическое взаимодействие с ней белков, узнающих определенные (например, регуляторные) последовательности нуклеотидов. В свою очередь, исследование взаимодействия вирусной РНК и белков может дать вклад в понимание механизмов процесса сборки и размножения вирусов, структурной организации и функционирования рибонуклеопротеидов.
Ниже изложена методика приготовления образцов для АСМ-исследования вирусной РНК, апробированная совместно с д. х. н. Ю. Ф. Дрыгиным (НИИ ФХБ им. А. Н. Белозерского МГУ).
Для приготовления всех растворов использовали деионизованную бидистиллированную воду. Препарат вируса (ВТМ, штамм U1), полученный методом дифференциального центрифугирования и центрифугирования в градиенте плотности раствора сахарозы, был любезно предоставлен Е. Н. Добровым (НИИ ФХБ МГУ).
В экспериментах по депротеинизации использовали суспензию ВТМ в воде при концентрации 2,6 мг/мл. Депротеинизацию вирусных частиц производили:
Для разделения отдельных молекул белковой оболочки ВТМ, свободной РНК и частично разрушенных вирусных частиц использовали метод высокоэффективной жидкостной хроматографии с гелем Toyo-Pearl HW-55. В трех используемых методиках колонки (150 мкл) предварительно уравновешивали водными растворами 36% ДМСО, 1,6 М мочевины или глицина (концентрация 0,1 М при pH 10,3), соответственно.
В качестве подложек использовали как чистые (свежий скол), так и модифицированные поверхности высокоориентированного пиролитического графита (пирографита) и слюды. Поверхность подложки модифицировали или катионным ПАВ - бензилдиметилалкиламмоний хлоридом (BAC) (1% раствор в формамиде, экспозиция 45 мин) [48] или 100 мМ MgCl2 (см. раздел 1.1.1), экспозиция 75 мин. Процесс модификации завершали промывкой водой (в 0,2 мл четыре раза по 1-2 мин), излишек воды удаляли фильтровальной бумагой и подложки высушивали в струе фильтрованного сжатого воздуха.
Молекулы РНК, высвобожденные из белковой оболочки ВТМ, или частично разрушенные вирусные частицы посредством обработки мочевиной или диметилсульфоксидом наносили на подложку из 1,6 М раствора мочевины или 36% диметилсульфоксида соответственно, разделив предварительно эти структуры на фракции по размерам методом высокоэффективной жидкостной хроматографии, см. выше.
Схема эксперимента без использования хроматографии включала следующие этапы. Вирусные частицы ВТМ (0,138 мг/мл) в 2 мМ MgCl2, 200 мМ KCl экспонировали 50% ДМСО и 3,7% формальдегидом в течении 5 мин при 45°С для денатурации РНК. Затем к смеси добавили 1% BAC и адсорбировали каплю препарата на поверхность слюды.
Молекулы РНК вируса энцефаломиокардита мышей, выделенные из вирусных частиц, иммобилизовали на подложке по следующей схеме. Макромолекулы (концентрация 60 мкг/мл) в 0,01 М HEPES буфере (pH 7,4) и 7,5% формальдегиде экспонировали в течении 15 мин при 55°C для денатурации. Затем или к препарату добавляли равный объем 0,1% раствора BAC в формамиде (до конечной концентрации BAC 0,05%) или к 3 частям препарата добавляли 27 частей того же раствора BAC (до конечной концентрации BAC 0,01%).
Адсорбцию исследуемых структур осуществляли (во всех случаях) из капли препарата (3-15 мкл), которую наносили на поверхность подложки и экспонировали во влажной среде под колпаком. Длительность процесса экспозиции варьировали от нескольких секунд до часа и более. После завершения экспозиции поверхность подложки также промывали водой и высушивали в струе фильтрованного сжатого воздуха.
АСМ-исследования проводили на воздухе в режимах постоянного или прерывистого контактов на приборе ``Nanoscope-IIIa'' (``Digital Instruments'', США). При проведении исследований в контактном режиме использовали кантилеверы жесткостью 0,06 Н/м из нитрида кремния. Для исследований в режиме прерывистого контакта применяли кремниевые кантилеверы с резонансной частотой в диапазоне 350-380 кГц, (параметры кантилеверов см. в таблице #.#).
Частицы вируса табачной мозаики были исследованы в режимах постоянного и прерывистого контактов на поверхностях пирографита и слюды. Было обнаружено, что скорость сорбирования вирусных частиц на свежий скол поверхности существенно выше при использовании пирографитовой подложки, чем при использовании подложки из слюды. В первом случае при равных условиях сорбирования (температура, объемная концентрация вирусов в препарате, время промывки и пр.) поверхностная плотность вирусных частиц существенно выше. В то же время на пирографитовой подложке при сканировании в режиме постоянного контакта вирусные частицы нестабильны и увлекаются иглой даже при минимизации воздействующей силы до величины менее 1 нН. Стабильные АСМ-изображения вирусных частиц на пирографите удается получить только в режиме прерывистого контакта. В случае же использования слюдяной подложки сила адгезии вирусных частиц достаточна для проведения исследований и в режиме постоянного контакта с величиной воздействующей силы в десятки нН.
Большая скорость сорбции вирусных частиц на поверхность пирографита в сравнении со слюдой может объясняться влиянием гидрофобного эффекта, см. раздел #.#.#;. В пользу данного утверждения говорит тот факт, что в случае гидрофобизации поверхности слюды посредством обработки BAC скорость сорбции вирусных частиц увеличивается.
Нестабильность адсорбированных на пирографит вирусных частиц в процессе сканирования в режиме постоянного контакта объясняется слабой адгезией частицы, обусловленной инертностью подложки (гидрофобный эффект не дает вклад в силу адгезии после высушивания образцов, поскольку проявляется только в водной среде).
АСМ-изображения частиц вируса табачной мозаики приведены на рис. 2.2. Согласно анализу полученных данных, для вирусных частиц среднее значение длины составляет 310 нм, при этом среднеквадратичное отклонение от среднего составляет 30%. Столь широкое распределение объясняется, во-первых, частичным разрушением вирусных структур при приготовлении препарата и, во-вторых, агрегацией вирусных частиц торец в торец (это, в частности, проявляется в наличии пиков на удвоенной, утроенной и т.д. длине вирусной частицы). С началом процесса обработки вирусных структур с целью высвобождения вирусной РНК максимум распределения вирусных частиц по длинам смещается в более короткую область.
| ,
Figure |
Анализ высот АСМ-изображений вирусных частиц над поверхностью подложки дает значение 18±2 нм (независимо от режима исследования), что согласуется с результатами электронной микроскопии [40]. Как показали дополнительные исследования, высота АСМ-профиля вирусной частицы над поверхностью подложки уменьшается при увеличении силы воздействия зонда, что связано с контактными деформациями, см. раздел #.#; результаты экспериментального исследования эффекта уменьшения высоты АСМ-изображений частиц ВТМ при увеличении силы воздействия изображены на рис. .
Частицы ВТМ, являясь природными ``наноразмерными'' объектами с фиксированными геометрическими параметрами, могут служить удобным тест-объектом для анализа качества зондирующего острия (определения параметров его геометрии). При минимизации силы воздействия зонда величина деформации вирусных частиц незначительна (см. экспериментальную зависимость рис. #.#; или рассчитанные значения деформации для модельных случаев, приведенные в таблице #.#). В силу этого можно считать, что частица вируса имеет в своем сечении окружность. Оценить эффект уширения (рассмотрению которого посвящен раздел #.#) для частицы, имеющей в сечении окружность, существенно проще, чем для частицы, имеющей эллипсоидальную форму (этот случай подробно рассматривается в разделе #.#). Согласно элементарным геометрическим построениям для острия с радиусом кривизны кончика R значение реально измеряемой ширины на полувысоте АСМ-изображений вирусных частиц радиуса r должно составлять величину:
| (2.1) |
Экспериментально определенные значения ширины АСМ-изображений вирусных частиц для режимов прерывистого и постоянного контактов были равны 30±5 нм и 32±6 нм соответственно. Воспользовавшись формулой (2.1), можно оценить радиус кривизны для кончиков зондов используемых типов кантилеверов: R ~ 5ё15 нм.
В мягких условиях депротеинизации наблюдали рибонуклеопротеиды (РНП) с выходящими из них (с одного конца или с обоих) нитями РНК, см. рис. 2.3 и 2.4. Мы протестировали три известные методики частичной депротеинизации ВТМ и высвобождения РНК, описанные в литературе [45,46,47]. Сравнительный анализ показал, что наиболее перспективным является метод с применением ДМСО (рис. 2.4). Метод депротеинизации в растворах со щелочным значением рН характеризуется большим фоном получаемых АСМ-изображений. Кроме того, молекулы РНК в этом случае склонны к агрегации и в значительной степени сохраняют элементы вторичной структуры. Метод с использованием мочевины позволяет получать АСМ-изображения с незначительным фоном, но при этом наблюдается агрегация вирусных РНП (рис. 2.3).
| ,
Figure |
| ,
Figure |
АСМ-изображения рисунков 2.3 и 2.4 получены в топографическом режиме Z(X,Y) и в режиме измерения сил трения Fтр(X,Y), (см. раздел #.#); последние изображения, в силу значительной разности высот частиц ВТМ и молекул РНК, характеризуются лучшим контрастом в сравнении с топографическими, позволяя различать на АСМ-изображении оба типа структур. Белковые остовы могут рассматриваться как маркеры молекул РНК, позволяя уверенно отождествлять наблюдаемые протяженные нити, выходящие из разрушенных вирусных структур, именно с молекулами вирусной РНК.
Наблюдение вирусных остовов с двумя нитями РНК, выходящими из одного конца вирусной частицы, может объясняться лишь процессом частичного их восстановления после того, как экспозиция с целью разрушения была завершена. Действительно, согласно работе [49], только в результате самосборки вирусной частицы возможно наблюдение двух нитей РНК, выходящих из вирусной частицы с одного конца, что связано с особенностью процесса самоорганизации вирусной частицы. Процесс самосборки вирусных частиц мог протекать только до разделения компонент препарата методом высокоэффективной жидкостной хроматографии, поскольку в процессе этой обработки молекулы белковой оболочки, молекулы свободной РНК и частично разрушенные вирусные структуры разделялись на отдельные фракции, что исключало возможность восстановления разрушенных вирусных частиц.
На заключительной стадии обработки вирусных частиц белковая оболочка разрушается, и макромолекула РНК полностью высвобождается. На рис. 2.5 приведены типичные результаты АСМ-исследования свободных РНК, адсорбированных на подложку из слюды.
| ,
Figure |
Молекулы, изображенные на рис. 2.5а и б, отделены от частично разрушенных вирусных частиц и молекул белковой оболочки методом высокоэффективной жидкостной хроматографии и адсорбированы на свежий скол слюды из 36% ДМСО; именно присутствие денатурирующего ДМСО способствует расправлению молекул.
Мы протестировали ряд методик модификации слюды до нанесения препарата: обработка в течении 75 мин 100 мМ MgCl2, 150 мМ KCl, 100 мМ MgCl2 + 150 мМ KCl; обработка 1% раствором BAC в формамиде или APTES (2 часа при 105° C), см. раздел 1.1.1. Было обнаружено, что методика с использованием MgCl2 приводит к компактизации молекул ДНК, адсорбируемых на подложке. Оптимальной, по-видимому, является методика с применением BAC; при этом хорошие результаты были получены и в случае, когда использовали свежий скол слюды, а BAC (конечная концентрация 0,01-1%) добавляли непосредственно к препарату, содержащему исследуемые макромолекулы. Таким образом, именно присутствие BAC способствует иммобилизации молекул на подложке.
Молекулы, изображенные на рис. 2.5в, также иммобилизованы на поверхности свежего скола слюды в присутствии 1% ВАС; методика приготовления препарата не включала в этом случае разделение его на фракции, поэтому для предотвращения самосборки частиц ВТМ молекулы РНК денатурировали обработкой 3,7% формальдегидом (см. описание экспериментальных методик 2.1).
Экспериментальный анализ ширины макромолекул РНК, адсорбированных на слюду, дает значения 10ё15 и 20ё30 нм для режимов прерывистого и постоянного контактов соответственно. Анализ значений высот макромолекул показал, что эти величины достаточно сильно варьируют как для различных молекул, так и для различных участков одной и той же молекулы, и заключены в диапазоне 0,3ё1,5 нм (для исследований как в режиме постоянного, так и прерывистого контактов). Данный разброс может объясняться сохранением макромолекулой элементов вторичной структуры (неполное разворачивание на поверхности подложки, наличие скрученных участков).
О частичном сохранении элементов вторичной структуры макромолекул говорит также наличие петель и ``шпилек''. Расправлению макромолекул может способствовать влияние потока при высыхании капли препарата (см. рис. 2.5). Это влияние обусловлено силами, действующими со стороны высыхающей капли, на молекулы, прикрепленные к подложке. Природа этих сил может быть гидродинамической или электростатической: действительно, капля препарата, находящаяся на поверхности слюды, обогащена ионами K+, перешедшими в раствор при частичной диссоциации слюды, поверхность молекулы РНК заряжена отрицательно. При высыхании капли концентрация катионов K+ увеличивается, что может вызвать электростатическое взаимодействие капли с молекулой РНК и объяснять их наблюдаемую ориентацию.
При этом на АСМ-изображении четко идентифицируются участки прикрепления макромолекулы к подложке. Видно, что молекула, будучи закрепленной в данной точке, ориентируется, расправляясь по потоку. Стабилизация макромолекул на поверхности подложки в расправленном состоянии позволяет провести анализ распределения макромолекул РНК по длинам. В то же время, если силы воздействия на молекулы значительны, то это может приводить к их разрыву, что способно затруднить интерпретацию получаемых распределений РНК по длинам, см. ниже.
На рисунке 2.6 представлена гистограмма распределения по длинам молекул РНК, высвобожденных из белковой оболочки посредством обработки ДМСО. Характер распределения одинаков как для молекул, адсорбированных из 36% ДМСО после разделения на фракции методом хроматографии, так и для молекул, адсорбированных в присутствии 1% BAC и денатурированных 3,7% формальдегидом (в обоих случаях адсорбция осуществлялась на свежий скол слюды). Из рисунка видно, что гистограмма распределения может быть описана суперпозицией трех гауссовых распределений (пунктирными линиями показаны соответствующие аппроксимирующие распределения, сплошной линией их сумма), характеризующихся значениями средней длины молекулы [`l]: 0,7 мкм, 1,4 мкм и 2,0 мкм. Значения ширины соответствующих распределений: 0,5 мкм, 0,3 мкм и 0,7 мкм соответственно.
|
Исследования зависимости длины высвобожденной РНК от длины частично разрушенного вирусного остова позволили сделать вывод, что полностью расправленным молекулам РНК соответствует пик со значением максимума 2,0 мкм. Поскольку РНК исследуемого штамма U1 включает около 6400 нуклеотидов, можно оценить величину межнуклеотидной связи в молекуле РНК, обработанной и иммобилизованной на подложку слюды по описанной методике: lн/н=0,31±0,02 нм.
С целью проверки универсальности апробированной при визуализации РНК ВТМ методики иммобилизации молекул на подложке с применением BAC, были проведены сходные исследования РНК вируса энцефаломиокардита (ВЭМК) мышей. Молекулы РНК, выделенные из препарата вируса, были денатурированы обработкой 7,5% формальдегидом в течение 15 мин при 55°C и адсорбированы на свежий скол слюды в присутствии 0,01 и 0,05% BAC, описание экспериментальных методик см. 2.1. Применение данного алгоритма позволило визуализировать молекулы исследуемой РНК в расправленном состоянии, см. рис. 2.7.
Этот результат позволяет сделать вывод о возможность иммобилизации однонитевой высокомолекулярной вирусной РНК (при условии ее денатурации) на поверхности слюды в расправленном состоянии по методике с использованием BAC; причем разработанный подход является, по-видимому, универсальным.
Иммобилизация РНК ВЭМК мышей на подложке в расправленном состоянии позволила провести анализ распределения их по длинам, см. рис. 2.8. При этом обнаружено, что характер распределения не зависит от используемой концентрации BAC.
|
Обнаружено, что распределение РНК ВЭМК мышей по длинам, изображенное на рис. 2.8 характеризуется несколькими максимумами, как и распределение по длинам молекул РНК ВТМ, приготовленных по сходной методике, см. рис. 2.6. Поэтому можно предположить, что специфический характер распределений обусловлен именно используемой методикой приготовления образцов.
Разработка методик иммобилизации вирусной РНК на поверхности подложки имела своей целью отработку рутинного подхода к исследованию специфического РНК/белкового взаимодействия; к картированию как данного взаимодействия, так и элементов вторичной структуры РНК с целью определения местоположения специфических последовательностей нуклеотидов. Тот факт, что молекулы РНК, иммобилизованные на подложке по апробированной методике, характеризуются несколькими максимумами в распределении по длинам, может быть учтен при решении задачи картирования следующим образом. Если построенная карта будет характеризоваться также несколькими максимумами, то это будет свидетельствовать о разной длине межнуклеотидной связи молекул, адсорбированных на подложке по используемой методике. В этом случае для определения адекватного местоположения исследуемой специфической последовательности нуклеотидов каждый максимум для расстояния от конца молекулы до специфического участка должен быть нормирован на соответствующий максимум для полной длины молекулы. Если же построенная карта будет характеризоваться единственным максимумом, то его следует нормировать на величину родительского максимума распределения молекул по длинам.
Молекула дезоксирибонуклеиновой кислоты является носителем генетической информации живых организмов (совокупности данных, необходимых для синтеза белков), которая ``записана'' в структуре молекулы в ``четвертичном'' (по числу нуклеотидов) коде. Согласно модели Уотсона и Крика [38] молекула ДНК представляет собой две нити, которые формируют двойную спираль, стабилизированную взаимодействием пар комплементарных нуклеодтидов (вторая нить образована нуклеотидами, комплементарными нуклеотидам первой нити).
В живой природе (внутри вирусов и клеток) молекула ДНК находится в существенно ``компактизованном'' состоянии, занимая объем на несколько порядков (до 106) меньший, чем объем молекулы ДНК в растворе (в хорошем растворителе). Таким образом, именно компактная конформация (конформация ``глобулы'') является ``естественной'' для молекулы ДНК, хотя в лабораторных исследованиях чаще имеют дело с ДНК в растворах, т.е. расправленной (конформация ``клубка'').
Перевод молекулы ДНК из развернутого в компактное (конденсированное) состояние может быть осуществлен посредством разнообразных конденсирующих агентов и пр.; структуру, образованную в результате конденсации ДНК в присутствии полимеров и солей, иногда называют psi-ДНК (psi расшифровывается как Polymer and Salt Induced) или y-ДНК [50]. Начало исследованию процессов компактизации ДНК было положено Лерманом [51], который методом седиментации обнаружил, что в разбавленных растворах ДНК, при увеличении концентрации нейтрального полимера (полиэтилен гликоля) и соли одновалентного металла (NaCl) выше некоторого порогового значения, формируются частицы высокой плотности.
В настоящее время установлено, что конденсацию молекул ДНК, находящихся в водных растворах, способны вызвать различные мультивалентные катионы: спермидин3+ [52], спермин4+ [53,54], катионные полипептиды: полилизин, гистоны [55], катионные ПАВ [56,57]. Двух- (и более) валентные металлические катионы способствуют конденсации молекул ДНК, находящихся в водно-спиртовых (этанол, метанол, изопропанол) средах [58].
Структура компактизованной ДНК исследовалась различными методами (краткий обзор основных результатов приводится в работе Блюмфельда [59]) при этом непосредственная визуализация объектов исследования осуществлялась методами электронной [55,60,61], атомно-силовой [62,63] и (с меньшим разрешением) флуоресцентной [56,57,64] микроскопии.
Результаты исследований показали [59], что двумя основными формами y-компактизованной молекулы являются тороидальная и стержневая. Для обеих форм сечение компактизованных структур, имеет, по-видимому, одно и то же значение (если структуры сформированы одинаковыми молекулами). Достаточно интригующим представляется то, что размер (радиус) тороидальной структуры не зависит от длины молекулы (если она лежит внутри диапазона 400- 40000 п.о.) и определяется, по-видимому, внутренними механизмами процесса конденсации; типичные значения внешнего радиуса тороидальной частицы Rout лежат в диапазоне 35-90 нм [59]. Длинные молекулы ДНК образуют компактные структуры из одной молекулы, при этом удавалось наблюдать тороидальную геометрию компактной частицы для молекул, длина которых не превышала 40000 п.о. Короткие ДНК формируют структуры, образованные в результате самоорганизации нескольких молекул, при этом совсем короткие молекулы (менее 150 п.о.), по-видимому, не могут быть компактизованы [65].
Метод электронной микроскопии, позволяя достигать высокого пространственного разрешения в латеральном направлении, дает значительную ошибку при определении высоты объекта, если она сравнима с размером зерна напыления; это обстоятельство не позволяет с большой точностью определять объем исследуемых частиц (необходимый для анализа их количественного молекулярного состава) по результатам исследований ЭМ. Более того, при расчете объемов частиц компактизованной ДНК часто не задаются целью определить высоту структур над подложкой, полагая, что сечение объекта представляет собой окружность [59]; нам представляется, что такое предположение не всегда обосновано (см. раздел 3.1.2).
Стоит также отметить, что специфика препарирования образцов для исследований ЭМ может искажать структуру объекта исследования.
Этого недостатка лишен метод флуоресцентной микроскопии, позволяющий проводить в реальном масштабе времени исследования компактизации молекул ДНК, находящихся в нативных условиях (в растворах с подходящим pH). Однако разрешающая способность метода ограничена пределом оптического микроскопа l/2. Поэтому в качестве объектов исследования приходится выбирать достаточно большие молекулы ДНК (например, гигантскую ДНК Т4, которая включает 166 тыс. п.о.) [56,57,64].
Применение метода АСМ для исследований компактизации молекул ДНК представляется весьма перспективным, поскольку оказывается возможным проводить исследования в нативных или близких к нативным условиях, при этом, как было показано в разделе #.#, геометрия исследуемых объектов может быть восстановлена путем анализа измеренных параметров их АСМ-изображений (высоты над подложкой и ширины на полувысоте). В то же время, разрешающая способность АСМ существенно выше, чем у метода флуоресцентной микроскопии и близка разрешающей способности ЭМ. Однако в настоящее время в литературе имеется лишь незначительное число публикаций [62,63], посвященных применению СЗМ для исследований процессов компактизации ДНК.
Авторы работы [62] исследовали начальные стадии конденсации плазмидной поли-А ДНК в результате ее взаимодействия со спермидином. Используемые концентрации спермидина были малыми (верхняя граница 150 мкМ) поскольку, как сообщают авторы, при больших концентрациях спермидин негативно влияет на возможность АСМ-визуализации исследуемых структур.
Эксперименты проводились на воздухе и адсорбция осуществлялась из капли препарата на поверхность свежего скола подложки (слюды). Авторы обнаружили, что при концентрации спермидина в диапазоне 7,5-15 мкМ отдельные участки ДНК образуют петли, радиус которых составляет величину около 20 нм. При больших концентрациях спермидина имеет место межмолекулярная агрегация ДНК, причем, в исследуемом диапазоне, с увеличением концентрации спермидина возрастают размеры агрегатов; специфическая форма этого класса структур оправдывает название ``цветка'', которое используют авторы для описания морфологии комплекса.
Авторы полагают, что наблюдаемые образования могут являться промежуточной стадией компактизации молекул в тороидальные структуры. Однако обращает на себя внимание специфическая плоская геометрия визуализованных образований: они характеризуются микронными значениями латерального размера при высоте всего 3-7 нм. Представляется, что в исследуемой системе весьма важен вклад влияния подложки на геометрию компактизованных комплексов, поэтому обнаруженная специфическая морфология может быть связана именно с тем, что изучаемая компактизация является, по сути, компактизацией на поверхности подложки; в этом случае естественно ожидать иной геометрии компактных структур, в сравнении с компактизацией в объеме.
Наиболее широко используемым способом иммобилизации молекул ДНК на подложке в расправленном состоянии 1.1.1 является метод добавления катионов металлов в рабочий раствор, содержащий нуклеиновые кислоты, с последующей адсорбцией капли препарата на поверхность слюды (свежий скол).
Типичные концентрации компонент в препарате, наносимом на поверхность слюды, обычно выбирают в следующем диапазоне: концентрация ДНК около 0,5 мкМ×п.о., концентрация MgCl2 или NiCl2 около 1 мМ. Каплю препарата объемом 0,5-5 мкл наносят на свежий скол слюды. На рис. 3.1а приведены результаты АСМ-исследования молекул ДНК бактериофага ФX-174 (5386 п.о.), которые были адсорбированы на слюду по описанной методике в присутствии NiCl2.
Другой метод фиксации макромолекул на подложке, это добавление к рабочему раствору, содержащему ДНК, молекул катионных ПАВ в малых концентрациях (менее 0,1%), см. рис. 3.1б. Типичные концентрации ДНК в препарате выбирают в том же диапазоне, что и выше. По-видимому, в этом случае имеет место комплексообразование молекул ДНК с молекулами ПАВ, образующими тонкую пленку, покрывающую снаружи каплю препарата, наносимую на подложку. Этот экспериментальный подход эквивалентен изложенному в разделе 2.1, где для иммобилизации вирусной РНК применяли метод комплексообразования молекулы нуклеиновой кислоты с ВАС, также являющимся катионным ПАВ.
Компактизация ДНК при взаимодействии с катионными ПАВ и прохождение комплексов ДНК-ПАВ через межфазную границу вода/малополярный органический растворитель может рассматриваться как отдаленная модель транспорта генетической информации в клетку [66].
Ранее процесс компактизации ДНК при взаимодействии с катионными ПАВ исследовался, в основном, лишь методом флуоресцентной микроскопии (ФМ) [56,57,64]. Это объясняется тем, что данный метод позволяет визуализировать только те объекты, которые ``окрашены'' молекулами флуоресцентного красителя, поэтому вклад во флуоресцентное изображение дает лишь молекула ДНК (именно с ней связываются молекулы красителя), а присутствие в растворе молекул ПАВ не влияет на получаемое изображение. В этом отличие и преимущество данного метода в сравнении с электронной и атомно-силовой микроскопией, для которых вопрос об адекватном распознавании и идентификации наблюдаемых структур может являться весьма нетривиальным. Этот аспект особенно существенен при исследованиях, связанных с молекулами ПАВ, в силу их способности к самоорганизации, в результате которой возможно формирование структур с различной геометрией [67].
Согласно результатам исследований авторов работы [28] свободная молекула ДНК характеризуется невысокой проводимостью при исследовании СТМ, что выражается в визуализации молекулы по способу отрицательного контраста (т.е. при движении над макромолекулой, в силу диэлектрических свойств последней, сканирующая игла опускается ниже, чем при движении над подложкой). Несмотря на это, мы обнаружили при исследовании комплексов ДНК-ПАВ, что последние могут быть успешно визуализованы методом СТМ (по способу прямого контраста).
Ниже изложена методика приготовления образцов для СЗМ-исследования комплексов ДНК-ПАВ, апробированная совместно с к.х.н. О. А. Пышкиной и к.х.н. В. Г. Сергеевым (Химический факультет МГУ).
В эксперименте использовали ДНК из эритроцитов цыплят (``Союзхимреактив'', Россия) 1000-3000 п.о. и дистеарилдиметиламмоний хлорид (ДСДАХ) (``Tokyo Kasei Kogyo Co.'', Япония).
Эксперименты проводили на СТМ СКАН-8 (``Центр перспективных технологий'', Россия) [68], в режиме постоянного тока (рабочие параметры: Ut ~ 0,3 В, It ~ 0,5 нА). АСМ-эксперименты проводили на приборе ``Nanoscope-IIIa'' (``Digital Instruments'', США) в режиме контакта с использованием кантилеверов из нитрида кремния (см. таблицу #.#).
Раствор ДНК (10-5 М) и ПАВ (9×10-5 М) в 0,5-М ТВЕ буфере объемом 2 мкл помещали на подложку (свежий скол пирографита или слюды) и высушивали в течение 3 мин. При нанесении комплексов из хлороформа их первоначально формировали в водной фазе при взаимодействии ДНК с ПАВ, затем комплексы высушивали и растворяли в хлороформе (в отличие от свободных молекул ДНК их комплексы с ПАВ растворимы в хлороформе [69]).
Было обнаружено, что комплексы ДНК-ПАВ могут быть визуализированы методом СТМ, см. рис. 3.2 а и б, где исследуемые объекты нанесены на подложку пирографита из водной фазы (а) и хлороформа (б). Для исключения эффекта влияния подложки и проверки адекватности применения метода СТМ к исследованию диэлектрических структур проводили контрольные измерения с помощью атомно-силового микроскопа. При этом были визуализованы объекты с идентичной морфологией (рис. 3.2 в).
Было обнаружено, что частицы комплекса ДНК-ПАВ хорошо адсорбируются на поверхность гидрофобной пирографитовой подложки из водной фазы, что может объясняться гидрофобным эффектом, см. #.#.# (поверхность исследуемых комплексов характеризуется гидрофобными свойствами [66]). Методика нанесения частиц комплекса из хлороформа позволяет избежать агрегации исследуемых структур в процессе приготовления образцов. Это может быть связанно с тем, что в хлороформе гидрофобное взаимодействие отсутствует и агрегировавшие образования реассоциируют.
Можно предположить, что возможность визуализации таких объектов, как комплексы ДНК-ПАВ, методом СТМ связана с поверхностной проводимостью слоя молекул ПАВ, покрывающих комплекс снаружи. Это предположение согласуется с обнаруженной нами ранее (см. работу [70]) возможностью СТМ-визуализации бислойных липидных образований - липосом. Действительно, исследованные липосомы были образованы молекулами липидов, свойства и структура которых близки молекулам используемого ПАВ (ДСДАХ).
В силу того, что комплексы ДНК-ПАВ нерастворимы в водной фазе, но растворимы в органических растворителях (хлороформе, гептане, циклогексане) [69], можно провести исследование процесса их переноса через границу раздела фаз в органический растворитель и исследовать морфологию перешедших комплексов. Подобная система может служить модельной при изучении механизма трансмембранного переноса ДНК [66].
Знание геометрии компактизованных комплексов является весьма важным, например, для определения состава комплекса (количества образующих его молекул). Точность определения геометрии органического объекта по результатам исследований электронной микроскопии ограничена (трудно получить точную информацию о высоте объекта: методика запыления под углом дает значительную ошибку при малых высотах объекта).
Определение геометрии комплексов ДНК-ПАВ по результатам относительных методов исследования (см. раздел 1.2) осложнено в силу вариации комплексов по размерам.
Метод АСМ позволяет непосредственно определять форму и размеры отдельных объектов, адсорбированных на поверхность подложки, но для восстановления их реальной геометрии необходим дополнительный математический анализ (учет эффекта уширения, см. раздел #.#). Здесь мы применили изложенную в разделе #.#; методику для определения формы комплексов ДНК-ПАВ, что позволило сделать выводы о количестве молекул ДНК, входящих в комплекс.
Ниже изложена методика приготовления образцов для АСМ-исследования комплексов ДНК-ПАВ, апробированная совместно с к.х.н. О. А. Пышкиной и к.х.н. В. Г. Сергеевым (Химический факультет МГУ).
Процесс переноса комплексов осуществляли из водной среды в хлороформ. В качестве ПАВ нами был выбран дистеарилдиметиламмоний хлорид (ДСДАХ) ``Tokio Kasei Kogyo Co.'', Япония. В работе использовали ДНК из спермы лосося (``ГОСНИИОХТ'', Россия) 300-500 п. о.
Свободные молекулы ДНК нерастворимы в хлороформе и могут быть переведены в органическую фазу только в виде комплексов с ПАВ (которые, как упоминалось, в хлороформе растворяются). Контроль процесса переноса комплексов ДНК-ПАВ через границу раздела фаз проводили методом УФ-спектрофотометрии на спектрофотометре ``Specord M40'' (Германия). Было показано, что при добавлении 1,5-кратного избытка ДСДАХ к двухфазной системе, состоящей из водного раствора ДНК и фазы хлороформа, наблюдается практически 100% перенос молекул ДНК в органическую фазу.
Адсорбцию образцов на слюду осуществляли стандартным образом: каплю препарата (хлороформ с комплексами после проведения 70% переноса) объемом 3 мкл помещали на свежий скол слюды, выдерживали в течение 3-5 минут, промывали бидистиллированной водой и высушивали на воздухе, после чего осуществляли сканирование.
АСМ исследования осуществляли на приборе ``Nanoscope-IIIa'' (Digital Instruments, США) в режиме контакта с кантилеверами из нитрида кремния жесткостью 0,06 Н/м.
Методика нанесения комплексов ДНК-ПАВ на поверхность подложки из хлороформа, по-видимому, имеет следующие преимущества в сравнении с нанесением из водных растворов. Комплексы ДНК-ПАВ являются гидрофобными, и в водной среде могут взаимодействовать с силами притяжения за счет гидрофобного эффекта, что может привести к агрегации комплексов при приготовлении образцов и осложнить их АСМ-визуализацию. В хлороформе гидрофобные взаимодействия не проявляются, и перешедшие туда комплексы стабилизированы лишь внутренними механизмами конденсации ДНК, в силу чего каждый комплекс, по-видимому, состоит из молекул ДНК, покрытых снаружи слоем молекул ПАВ. Капля хлороформа, содержащая исследуемые комплексы, при нанесении на подложку высыхает достаточно быстро, что уменьшает вероятность образования регулярных структур вследствие механизмов агрегации или самоорганизации ПАВ.
С помощью АСМ было обнаружено, что при нанесении на подложку структур, оставшихся в водной фазы после проведения 70%-переноса, на АСМ-изображениях присутствуют агрегаты молекул ДНК в виде нитей длины 250-2000 нм. Образование длинных тяжей происходит, по всей видимости, вследствие межмолекулярной агрегации отдельных молекул ДНК при воздействии ПАВ. Измерение значений высоты и ширины на полувысоте АСМ-изображений данных структур дает значение 0,3-0,9 нм и 13-25 нм соответственно.
На рис. 3.3 приведены результаты АСМ-исследования комплексов ДНК-ПАВ, перешедших в хлороформ, адсорбированных затем на свежий скол слюды и исследованных на воздухе. С целью анализа морфологии комплексов была проведена статистическая обработка измеренных параметров АСМ-изображений тороподобных структур: D - диаметра тора, d - диаметра сечения тора и h - высоты тороидальной структуры над подложкой.
|
Гистограммы распределения параметров тороидальных комплексов представлены на рис. 3.4, статистика набрана в результате измерения 180 АСМ-профилей комплексов, сплошной линией указана аппроксимация полученных распределений по закону распределения Гаусса.
|
Средние значения для параметров тора, определенные по аппроксимации распределением Гаусса, составляют следующие величины (в качестве погрешности указано стандартное отклонение):
D = 100±30 нм, d = 25±9 нм и h = 5±2 нм.
Т.о. параметры частиц (внешний радиус тора Rout = (D+d)/2 = 62±16 нм) хорошо согласуются с типичными значениями для молекул ДНК, имеющих различную длину и сконденсированных различными агентами [59].
В силу того, что длина молекул лежит в диапазоне 100-170 нм, а средняя длина окружности тороидальной частицы pD составляет величину 300 нм, очевидно, что в образовании комплекса принимает участие, по крайней мере, две-три молекулы ДНК. Более подробный анализ требует восстановления реальной геометрии комплекса.
Занижение значений вертикальных размеров исследуемых структур обусловлено контактными деформациями под действием зонда, особенно существенными при исследовании объектов, радиус кривизны которых меньше радиуса кривизны кончика иглы, см. раздел #.#. Предполагая, что деформация объекта не сопровождается изменением его объема, мы делаем вывод о том, что уменьшение высоты АСМ-изображения (занижение параметра h = 2b) будет скомпенсировано соответствующим увеличением его ширины (параметра d). Таким образом, мы делаем априорное предположение, что тороидальные комплексы ДНК-ПАВ имеют ``сплюснутую'' форму.
Завышение значений ширины АСМ-изображений является в основном следствием того, что кончик зондирующего острия имеет конечный радиус кривизны, см. раздел #.#. Рассчитать вклад в уширение можно на основании анализа геометрии контакта исследуемой структуры и кончика зонда с помощью программы .
Результаты применения методики учета эффекта уширения (раздел ) к восстановлению ширины сечения исследуемых объектов.
Геометрию кончика иглы (радиус кривизны) можно определить при исследовании объектов, имеющих в сечении окружность известного радиуса, см. 2.1. Согласно результатам исследования тест-объектов (частиц вируса табачной мозаики, см. раздел 2.1) радиус кривизны кончика иглы кантилеверов используемого типа имеет величину от 5 до 15 нм (по данным фирмы-производителя до 40 нм, см. таблицу #.#). Определение точного значения радиуса R для конкретного зонда требует его тестирования непосредственно перед использованием. Однако и в этом случае существует вероятность того, что в процессе сканирования форма иглы претерпит изменения в результате взаимодействия с объектом. В этой связи чрезвычайно полезным является получение информации о форме зонда непосредственно из АСМ-изображений объекта исследования. Эта информация может быть получена в методике учета эффекта уширения (раздел ).
Так, на рис. 3.5 представлен график зависимости числа случаев отсутствия решения (для статистики пар значений d и h) от радиуса аппроксимации зонда, построенный путем тестирования выполнения ``реперных'' условий () или (). На основании рис. 3.5 можно сделать вывод, что верхняя граница значения R, характеризующего используемый зонд, составляет 12 нм - выше этого значения наблюдается линейный рост числа случаев отсутствия решения. Нижняя граница для радиуса кривизны зонда определяется контактными деформациями.
|
Восстановление истинной ширины тороидальных структур дает среднее значение параметра a: 10-11 нм (относительное отклонение e = 0,5) в моделии сферической иглы. Результаты применения параболической модели иглы дают значения 11-12 нм. Таким образом, действительно, формой комплекса ДНК-ПАВ является сплюснутый тор.
Расчет числа молекул ДНК, образующих комплекс с ПАВ.
Предполагая плотную (гексагональную) упаковку молекул ДНК в комплексе, получим, что объем, приходящийся на одну молекулу в составе комплекса, имеет величину 4 R2 sin60° L, где R - радиус молекулы ДНК, а L - ее длина.
Объем тороидального комплекса будет определяться формулой p2 abD, где a - найденное численное решение, b = h/2, D - диаметр тора. С учетом наличия слоя ПАВ толщиной l получим формулу для числа молекул:
|
На рис. 3.6 приведены гистограммы распределения найденных значений числа молекул при двух значениях параметра R (радиуса кривизны иглы) - 6 и 12 нм.
|
Анализ гистограмм показывает, что, согласно полученному решению, 80% комплексов имеет в своем составе менее 15 молекул ДНК (при R=12 нм).
В разделе 3.1 мы рассматривали конформационные изменения, которые претерпевают молекулы ДНК при взаимодействии с молекулами ПАВ в водных растворах. Методом АСМ было подтверждено, что в этих условиях может происходить компактизация молекул ДНК, при этом была визуализирована тороидальная структура компактных комплексов.
Стоит отметить, что система ДНК + ПАВ достаточно сложна для исследований прямыми методами визуализации геометрии микрообъектов, к которым относится и АСМ. Действительно, присутствие ПАВ в качестве компонента системы требует проведения трудоемких контрольных экспериментов для определения методики приготовления образцов, при использовании которой структуры, образованные только ПАВ (без ДНК), не приводили бы к ошибкам в интерпретации АСМ-результатов. Это связано со способностью ПАВ к самоорганизации [71], которая, например, может иметь место в процессе высыхания капли препарата, нанесенной на поверхность подложки для приготовления образцов (при высыхании капли будет повышаться концентрация ПАВ). Поэтому необходимо определить такие условия процессов нанесения образцов, высушивания капли препарата и промывки, при которых наличие на подложке объектов, связанных лишь с самоорганизацией ПАВ (без участия ДНК), минимально. Или, если они и присутствуют, то их геометрия принципиально отличается от исследуемых комплексов ДНК-ПАВ: например, наличие на подложке самоорганизованных плоских моно- или мультислойных пленок ПАВ не осложняет идентификацию искомых ДНК-ПАВ комплексов, но, напротив, часто способствует иммоблизации исследуемых объектов в процессе сканирования.
Поэтому достаточно показательным было бы подтвердить адекватность применения СЗМ для исследования процессов конденсации ДНК на примере более простых систем. Таковой системой, на наш взгляд, может считаться молекула ДНК, находящаяся в водно-спиртовой среде (этанол, изопропанол). При достаточно высоких концентрациях спирт становится для молекул ДНК плохим растворителем (в присутствии катионов Na+ - при работе с натриевыми солями ДНК), что приводит к переходу макромолекул из состояния развернутого клубка в состояние компактной глобулы [72].
Ниже изложены методики приготовления образцов для АСМ-исследования конденсации молекул ДНК, апробированные совместно с к.х.н. О. А. Пышкиной и к.х.н. В. Г. Сергеевым (Химический факультет МГУ).
Эксперименты в естественной атмосфере осуществляли по следующей схеме. Молекулы ДНК конденсировали в водно-спиртовой среде при концентрациях спирта, приводящих к переходу клубок ® глобула, и наносили каплю препарата на подложку, затем образцы высушивали, промывали и снова высушивали, т.е. схема эксперимента была стандартной для исследований на воздухе и идентичной случаю анализа конденсации комплексов ДНК-ПАВ.
Для экспериментов выбрали ДНК из эритроцитов цыплят (1000-3000 п. о., ``Союзхимреактив'', Россия ), в качестве спирта использовали изопропанол и этанол (х. ч.). Контрольные эксперименты по исследованию молекул ДНК, приготовленных нанесением из водного раствора буфера (без спирта), демонстрируют отсутствие компактизации макромолекул в данных условиях - рис. 3.7. Здесь мы использовали в качестве подложки немодифицированную слюду, поскольку такую же подложку выбирали и для конденсированных молекул. Использование немодифицированной (т.е. отрицательно заряженной) слюды в качестве подложки объясняет тот факт, что адсорбированные молекулы недостаточно расправлены и частично проагрегировали.
|
Добавление к исходному раствору 50% и более спирта (изопропанола или этанола) существенно влияет на конформацию ДНК. Результаты экспериментов показывают, что большая часть молекул находится в одной из двух типов конформаций - тороидальной или стержневой (rod-like). Следует подчеркнуть, что при высокой концентрации молекул ДНК в исходном растворе тороидальные и стержневые частицы агрегируют в единую структуру. На рис. 3.8 представлен результат АСМ визуализации стержневой структуры ДНК, полученной при адсорбции макромолекул из 66% водного раствора этанола (концентрация ДНК составляла 5×10-5 М).
|
Анализ ширины профиля стержневых структур показывает, что они образованы, по крайней мере, несколькими молекулами ДНК (``стянутыми в жгут''); высокая степень агрегации стержневых структур объясняется значительной концентрацией ДНК в рабочем растворе.
Несколько отличные результаты были получены при использовании изопропанола. На рис. 3.9 представлены результаты АСМ-визуализации тороидальных структур, образованных молекулами ДНК при нанесении на поверхность слюды 4×10-7 М раствора ДНК в 50% изопропаноле (в присутствии 0,05 М ТВЕ буфера). Результаты свидетельствуют, что в этом случае (в сравнении с этанолом) более часто наблюдаются тороподобные структуры, нежели стержневые. Причина может заключаться в различных значениях диэлектрических проницаемостей изопропанола (e = 18,3) и этанола (e = 24,3), что обеспечивает лучшую компенсацию заряда поверхности макромолекулы противоионами Na+ в случае изопропанола.
|
Стоит подчеркнуть, что при проведении АСМ-исследований на воздухе необходимой стадией приготовления образцов является процедура высушивания капли препарата на подложке. В процессе высушивания концентрации компонент препарата в капле неравновесно возрастают, что может неконтролируемо влиять на морфологию адсорбируемых на подложку структур. Исключить процедуру высушивания возможно при проведении АСМ-исследований в жидких средах.
В этой серии экспериментов проводили исследования конденсации молекул ДНК непосредственно в жидкостной ячейке АСМ, что позволяло исключить возможное негативное влияние процесса высушивания капли препарата в стандартной схеме экспериментов на воздухе.
Использовали ДНК бактериофага Т4 и водные растворы изопропанола и этанола. Ранее было показано [72], что гигантская ДНК Т4 в водно-спиртовой смеси при концентрации спирта более 50% (для изопропанола) претерпевает конформационный переход клубок ® глобула. При уменьшении концентрации спирта до 40% наблюдалось частичное разворачивание глобул. Динамика перехода исследовалась методом флуоресцентной микроскопии. Однако, в силу ограничения разрешающей способности метода (предел оптического микроскопа), оказалось трудным определить точную геометрию компактных глобул и структуру частично компактизованных образований.
Представлялось показательным провести АСМ-исследования процесса компактизации именно для этого объекта в тех же условиях.
В качестве объектов исследования использовали ДНК бактериофага Т4 (``Nippone Gen'', Япония), состоящей из 166 тыс. п.о. При проведении экспериментов использовали бидистиллированную воду, очищенную с помощью системы ``Milli-Q'' (``Millipore'', США). Водно-спиртовые среды готовили с использованием изопропанола (х. ч.).
Исследования конденсации молекул ДНК проводили методом атомно-силовой микроскопии в водно-спиртовой смеси с использованием подложек слюды. Свежий скол слюды и кантилевер (с зондом) помещали в жидкостную ячейку АСМ, которую последовательно заполняли рабочими растворами. Препарат, содержащий молекулы ДНК Т4 в водно-спиртовой смеси при 80% концентрации спирта, получали, смешивая один объем раствора ДНК в 0,5-ТВЕ-буфере с четырьмя объемами изопропанола. Препарат молекул ДНК в 40% изопропаноле получали, смешивая два объема раствора ДНК в буфере с тремя объемами изопропанола. В обоих случаях конечная концентрация ДНК в смеси составляла 1×10-6 М.
Эксперименты проводили в жидкостной ячейке АСМ ``Nanoscope-III'' (Digital Instruments, США) в режиме прерывистого контакта с использованием заостренных кантилеверов из нитрида кремния жесткостью 0,32 Н/м, см. таблицу #.#.
Определение адекватной методики проведения АСМ-эксперимента включало анализ наиболее подходящих концентраций водно-спиртовой смеси, а также последовательность заполнения жидкостной ячейки рабочими растворами (тестировали различные варианты).
В результате тестирования остановились на следующей методике:
Препарат, содержащий молекулы ДНК Т4 в водно-спиртовой смеси при 80% концентрации изопропанола, вводили в жидкостную ячейку, заполненную предварительно водно-спиртовой смесью с той же концентрацией спирта. При этих условиях макромолекулы ДНК осаждались на поверхность слюды в виде компактных глобул (рис. 3.10а и в).
| , Figure |
Применение методики восстановления реальных размеров исследуемых объектов по измеренной высоте и ширине профиля АСМ-изображения (учет эффекта уширения), которая изложена в разделе 2.2, позволяет определить объем глобулярных структур. Проводили расчеты эффекта уширения для двух значений радиуса кривизны иглы: 5 и 10 нм, результаты приведены в таблице 3.1. Согласно оценкам (с применением тест-объектов) радиус кривизны иглы, используемой при записи приведенных изображений, лежит именно в диапазоне 5ё10 нм. Значения параметров глобулы, определенные с использованием двух граничных значений для радиуса кривизны иглы фактически идентичны. Восстановленные значения параметров полуосей a и b позволяют сделать вывод, что геометрической формой глобулы является сплюснутый и слегка вытянутый эллипсоид.
Стандартные отклонения величин, приведенных в таблице 3.1, велики, что объясняется статистическим разбросом измеряемых параметров для различных глобул. Более наглядно это видно из анализа гистограмм распределения анализируемых параметров.
| R, нм | a, нм | b, нм | c, нм | V, ×105 нм3 |
| 5 | 41±12 | 55±17 | 23±9 | 2,5±1,7 |
| 10 | 37±12 | 52±17 | 23±9 | 2,1±1,5 |
| ,
Figure |
На рисунке 3.11 представлены гистограммы распределения значений объема глобулярных частиц. Наличие в набранной статистике глобул, характеризующихся малыми значениями объема (меньше, чем объем одной молекулы ДНК) объясняется, по-видимому, присутствием в используемом препарате фрагментов молекул ДНК. Из анализа рисунка 3.11 (ср. с таблицей 3.1) следует, что объем глобулы, образованной цельной молекулой ДНК (не фрагментом) составляет величину порядка 3-5×105 нм3, что превышает величину объема молекулы ДНК Т4: 1,7×105 нм3.
Т.о. образом можно предположить, что каждая глобула образована одной молекулой ДНК, находящейся в достаточно плотно упакованном состоянии.
Частично декомпактизованные структуры макромолекул ДНК Т4, получаемые при разбавлении 80% спиртового раствора в воде до 40-50%, исследовали следующим образом. В жидкостную ячейку, содержащую ДНК Т4 в виде глобул в 80% изопропаноле (20% воды), добавляли смесь изопропанола с водой (40% изопропанола + 60% воды), с целью понизить концентрацию спирта в ячейке.
Поскольку процесс компактизации ДНК в спирте обратим, то при понижении концентрации спирта в жидкостной ячейке глобулы претерпевают конформационные изменения, связанные с началом процесса декомпактизации. На рисунках 3.10 б и г изображены те же участки поверхности, что и на рисунках 3.10 а и в, но при понижении концентрации спирта в ячейке до значения 40-50%. Из рисунков видно, что в процессе декомпактизации изменились очертания глобулярных структур и их высота (ср. а и б).
Некоторые выводы о структуре компактизованной глобулы можно сделать, анализируя стадии динамического процесса декомпактизации, изображенные на рис. 3.10 в и г. Анализ рисунка показывает, что первоначально компактная глобула (рис. 3.10 в, отметка ``Гл'' ) в процессе декомпактизации уменьшается в высоте и позволяет визуализировать центральную полость, см. рис. 3.10 г и врезку.
Однако оказалось, что дальнейший процесс разворачивания глобул не происходит. Это можно объяснить наличием сил взаимодействия между молекулой и подложкой, которые и препятствуют разворачиванию при уменьшении концентрации спирта. (Оказалось, что в условиях водно-спиртовой среды исследуемые объекты достаточно стабильны на подложке и не увлекаются зондом при сканировании).
Поэтому промежуточные состояния компактизации ДНК (между глобулой и клубком) получали следующим образом. Молекулы ДНК, находящиеся в 40% изопропаноле, вводили в жидкостную ячейку, а затем концентрацию спирта в ячейке повышали прокачиванием 80% изопропанола. При повышении концентрации спирта молекулы выпадали на поверхность подложки в частично компактизованном состоянии. Дальнейшая компактизация ДНК, адсорбированных на подложку, не наблюдалась, что, по-видимому, опять же объясняется взаимодействием молекул ДНК со слюдой.
АСМ-изображения частично компактизованных структур приведены на рис. 3.12. Было обнаружено, что начальным процессом компактизации глобул является закручивание отдельных участков макромолекулы ДНК в тороидальную структуру, эти компактные участки, по-видимому, и являются центрами дальнейшей компактизации.
| ,
Figure |
На основании наблюдений можно предположить, что и компактные глобулы, визуализованные на рисунке 3.10 а и в являются продуктом именно тороидальной компактизации макромолекул ДНК и представляют собой плотные ``клубки'', в которых отдельные участки молекулы закручены вокруг тороидальных структур.
Следует отметить, что макромолекулы ДНК при повышении концентрации спирта в ячейке (от 40%) образуют не только тороидальные, но и (реже) стержнеобразные структуры, см. рисунок 3.13.
| |
Стрежневые структуры, изображенные на рисунке 3.13, имеют ту же природу, что и приведенные на рисунке 3.8, где изображены результаты исследований, проводимых на воздухе. Видно, что при исследованиях в жидкости достижимо большее пространственное разрешение, что позволяет визуализировать отдельные нити молекулы ДНК, образующей исследуемую структуру. Из рисунка 3.13 б видно, что при сканировании под воздействием зонда микроскопа стержневая структура частично ``расплелась''.
Наблюдение в ряде случаев компактизованных структур, имеющих морфологию, отличную от тороидальной, может объясняться тем, что в эксперименте исследовались промежуточные стадии процесса компактизации, в которых некоторое количество молекул может находиться в нестационарных морфологических состояниях
Описанные подходы проведения АСМ-эксперимента в жидких средах (т.е., потенциально, в условиях, близких нативным) могут быть полезны при исследованиях модельных систем транспорта генетического материала внутрь живых клеток. Подобные исследования могут также дать вклад в понимание механизмов процессов компактизации молекулы ДНК под воздействием различных компактизующих реагентов.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
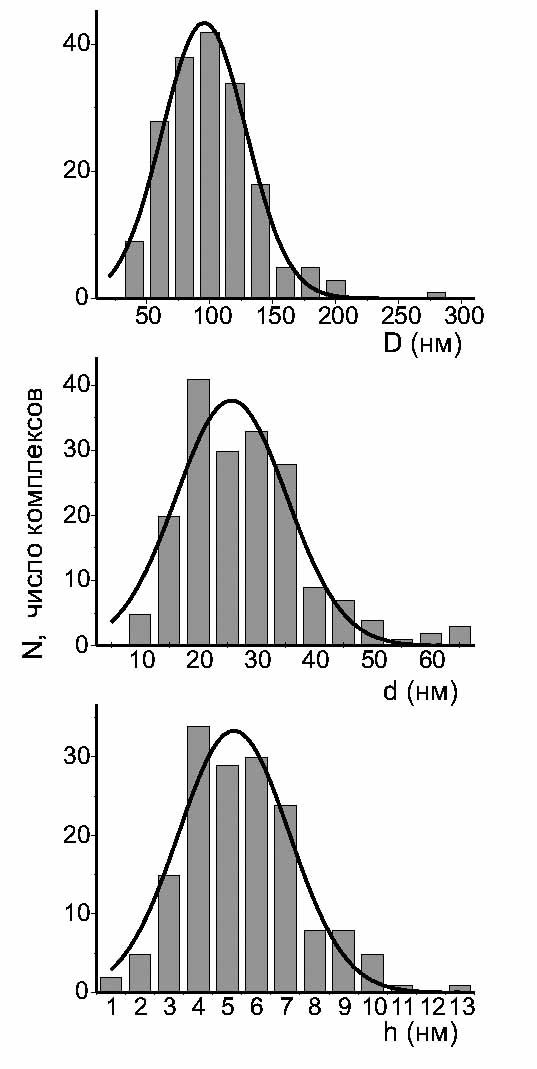{kind=link}
{kind=link}
{kind=link}
{kind=link}
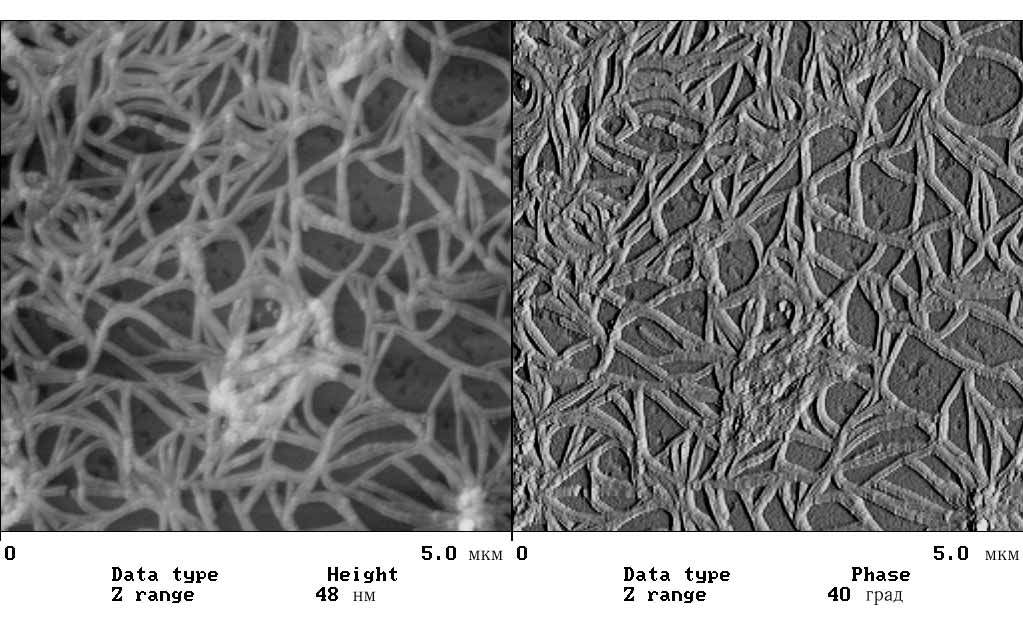{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}